![]()

The Sangiran Homo erectus locality is located near the village of Sangiran, central Java, Indonesia. Fossils were first collected from this site, on behalf of Ralph von Koenigswald, by local villages between 1936-1940 (von Koenigswald, 1938,1940). Collection was suspended during WWII but continued after 1950 by farmers and Indonesian researchers (Sartono, 1968, 1971; Jacob, 1966). Currently approximately 80 hominin fossils, including 10 partial crania and 14 mandibular/maxillary specimens have been catalogued from the site. The fossil collected by von Koenigswald are held in the Senckenberg Forschungsintitut in Frankfurt, Germany, with the post WWII collections held at the GRDC in Bandung and Gadjah Mada University, Yogyakarta.
Fossil hominin bearing formations of the Sangiran dome are the Sangiran formation and the overlying Bapang formation. These two formations are separated by a fossil-rich layer, which used to be called the Grenzbank zone. While the stratigraphic integrity and dating of the hominin fossils remains somewhat problematic, the oldest fossils apparently derive from the top of the Sangiran formation. However, most of the hominin fossils come from the younger Bapang formation. Based on Argon-Argon dating the Sangiran formation dates to the period 1.92-1.58 myr and Bapang 1.58-1.0 myr. The Notuporo formation that caps the Bapang, and has an Argon-Argon date of 780 ka is currently without hominin fossils, so the geologically youngest fossils at Sangiran may be approximately 1 myr in age. Uncertainties continue over to what extent the hominin fossils are secondary depositions in a reworked deposit and the manner in which the fossils have been collected complicates the identification of their precise stratigraphic contexts.
![]()
 |
 |
|---|---|
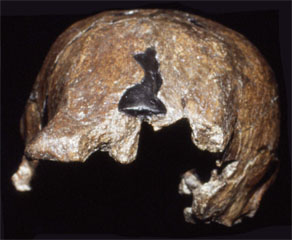
| Sangiran 2 | Sangiran 2 |
 |
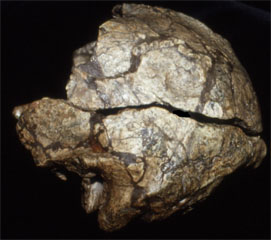 |
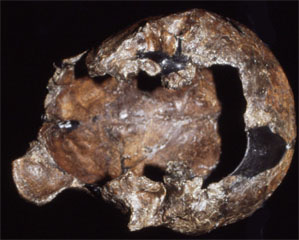
| Sangiran 2 | Sangiran 3 |
The first partial cranium of H. erectus (Sangiran 2) was discovered in 1937, with Sangiran 4 (partial cranium with palate and most of the maxillary teeth) found in 1939. The most complete specimen so far discovered is Sangiran 17 and this is curated at the GRDC. All of the Sangiran hominin fossils are now allocated to Homo erectus (formerly Pithecanthropus and Meganthropus) Until recently it was thought that H. erectus was the first hominin to have dispersed from Africa, and to have subsequently spread as far as northern China (Zhoukoudian) and Java. Many researchers also thought that H. erectus populations were likely ancestral to H. sapiens and this was incorporated in the multiregional model of human evolution (Thorne and Wolpoff 1981). However, based on current interpretations of the anatomy of Javan H. erectus (Kaifu et al., 2005, 2008; Baba et al., 2004; Brown 2009) the absence of early evidence for H. sapiens from the Indonesian archipelago (Brown 2012), and the assessment of evolutionary relationships provided by DNA (Rasmussen et al., 2011) it is probable that Javan H. erectus became extinct in the middle-late Pleistocene. Modern humans (H. sapiens) may not have been present in southeast Asia until after the Toba super eruption of 69-77,000 years BP. This eruption may also have contributed to the demise of Javan H. erectus. However, the volcanic plume spread primarily to the northeast of Sumatra, towards the Bay of Bengal and India, and there is no convincing evidence of associated loss of fauna and vegetation in southeast Asia (Louys, 2007). This will be discussed further in the entries on Sambungmacan and Ngandong.
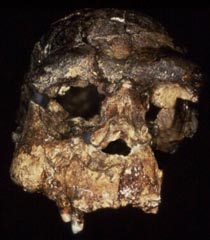
Sangiran 17, discovered in 1969 was initially described by Sartono (1973) and referred to as Pithecanthropus VIII. Both Sartono (1975) and Jacob (1975) highlighted anatomical similarities with the Ngandong hominins (Weidenreich 1951; Santa Luca 1980) and thought that it should be placed in the same taxon. With an endocranial volume of 1029-1125 cc, Sangiran shares a distinctive set of anatomical features with the other Javan H. erectus from Sangiran, Ngandong and Sambungmacan. The The cranial vault is long and low, with a receeding frontal bone and sharp angulation of the superior and inferior nucal planes. The supraorbital torus form a relatively straight and thickened bar, which is particularly thikened laterally. The frontal bone has a median ridge and and bone surface is raised and thickened near bregma. There is marked postorbital constriction and the superior temporal lines are pronounced and relatively high on the sides of the vault. Maximum cranial breadth is located on the supramastoid crests. While the cranial vault bone is thickened, it differs from the pattern in H. sapiens, primarily with thickened bone in the lateral cranial walls and in the cranial base.There is a pronounced occipital torus and rugose areas of neck muscle attachment. The facial skeleton has some postmortmem crushing and distortion and can not be perfectly articulated with the frontal bone. While facial prognathism is pronounced, it is less than in the original attempts at reconstruction. The orbits are deep and relatively circular in shape, and the nasal aperture does not appear to have been very broad. The most distinctive aspect of the face is in the extremely deep, thickened and strengthened, preserved right zygomatic bone.The inferior margin of this bone merges directly with the tooth row. Areas of masticatory muscle attachment and structural reinforcement suggest an adaptation for forceful chewing, however, the maxillary teeth are surprisingly small and with minimal occulsal wear for a hunter gatherer. The Sangiran mandibles also combine thickened tooth bearing segments (corpus and alveolar bone), with surprisingly small teeth.
Thorne and Wolpoff (1981) argued that the morphology of Sangiran 17, particularly in the details of fronto-facial morphology, prognathism, vault bone thickness and tooth-size, were indicative of a regional anatomical pattern that demonstrated evolutionary continuity between Javan H. erectus and late Pleistocene H. sapiens in Australia. However, their reconstruction of Sangiran 17 greatly exaggerated its facial prognathism (Aziz et al., 1996) and neither Pleistocene or more recent Australian Aboriginal crania have the distinctive combination of anatomical traits identified in Javan H. erectus (Groves, 1989; Lahr, 1996; Stringer, 1998; Brown 2010). An ancestor-descendent relationship between Javan H. erectus and Australian Aborigines is also dismissed by analyses of DNA from both recent humans and the Pleistocene Denisovians (Rasmussen, et al., 2011; Reich, et al., 2011).
 |
 |
|---|---|
| Sangiran 17 | Sangiran 17 |
 |
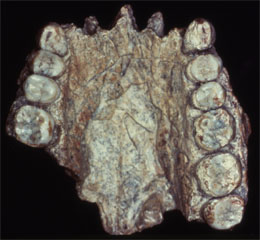 |
| Sangiran 27 | Sangiran 4 |
![]()
 |
craniometric data for LB1 H. floresiensis, H. erectus and H. habilis |
|---|
![]()
References
Brown, P., 2010. Nacurrie 1: Mark of ancient Java, or a caring mother’s hands, in terminal Pleistocene Australia? J. Hum. Evol. 59, 168-187.
Chen, C.H., Lee, M.Y., Yang, T.F., Iizuka, Y., Wei, K.Y. 2000. Newly discovered eastern dispersal of the youngest Toba Tuff. Marine Geology 167: 303-312.
Groves, C.P., 1989. A Regional Approach to the Probelms of the Origin of Modern Humans in Australasia., in: Mellars, P., Stringer, C. (Eds.), The Human Revolution. Behavioural and Biological Perspectives on the Origins of Modern Humans. Princeton University Press, Princeton, New Jersey.
Kaifu, Y., Aziz, F., Indriati, E., Jacob, T., Kurniawan, I., Baba, H., 2008. Cranial morphology of Javanese Homo erectus: New evidence for continuous evolution, specialization, and terminal extinction, J. Hum. Evol., pp. 551-580.
Lahr, M.M., 1996. The evolution of modern human diversity. Cambridge University Press, Cambridge.
Louys, J., 2007. Limited effect of the Quaternary's largest super-eruption (Toba) on land mammals from Southeast Asia. Quaternary Science Reviews, 26, 3108-3117.
Rasmussen, M., et al., 2011. An Aboriginal Australian Genome Reveals Separate Human Dispersals into Asia. Science 334, 94-98.
Reich, D., et al.., 2011. Denisova Admixture and the First Modern Human Dispersals into Southeast Asia and Oceania. The American Journal of Human Genetics 89, 516-528.
Stringer, C., 1998. A Metrical Study of the Wlh-50 Calvaria. J. Hum. Evol. 34, 327-332.
Swisher, C.C., Curtis, T., Jacob, T., Getty, A.G., Suprijo, A., Widiasmoro, 1994. Age of the earliest known hominid in Java, Indonesia. Science 263, 1118-1121.
Thorne, A.G., Wolpoff, M.H., 1981. Regional continuity in Australian Pleistocene hominid evolution. Am. J. Phys. Anthropol. 55, 337-349.
von Koenigswald, G.H.R., 1938. ein neuer Pithecanthropus-Schädel. Proceedings of the Academy of Science Amsterdam 41,185-192.
Weidenreich, F., 1951. Morphology of Solo Man. Anthrop. Pap. Am. Mus. Nat. Hist. 43, 205-290.
![]()